
Saludos a todos.
En esta ocasión, hablaremos de algunos datos curiosos sobre
los terópodos que usualmente, no se mencionan en los medios o bien, éstos
presentan de forma errónea o hasta contradictoria. Pues es de tener en cuenta
que en las películas, videojuegos e incluso, en algunos documentales, se suele mostrar
una imagen errónea sobre estos animales de modo que ésta resulte ser más
cautivadora para el público, llevando a percepciones que a menudo son
inconsistentes con lo que nos revela la ciencia. Y antes de comenzar, avisamos
que algunos de estos puntos pueden parecer desconcertantes para algunas
personas (de ahí, el título de la entrada) y por esta razón, advertimos que
proceda con precaución y moderación al leer el resto de la misma. Dicho esto, continúe
bajo su discreción.
Muy bien, comencemos:
1) Muchos terópodos poseen plumas
Si está familiarizado con este grupo de dinosaurios, este
dato no le resultará nada sorprendente. Y es que durante años, se han
descubierto pruebas de que numerosas especies de terópodos poseían algún u otro
tipo de plumaje en su cuerpo. Generalmente, las especies en las que se ha
encontrado evidencia de plumaje pertenecen a un grupo de terópodos conocido
como los coelurosaurios (Coelurosauria), el cual incluye, por mencionar algunos
ejemplos, al linaje del Velociraptor (los deinonycosaurios), al del
Tiranosaurio (los tiranosauroides) e incluso, al de las aves (Avialae). Así es,
las aves también son terópodos.
Esto nos dice que el ancestro común de estos dinosaurios
probablemente también poseía algún tipo de plumaje, ya que desde la perspectiva
evolutiva, es muy poco probable que este tipo de estructura se haya
desarrollado de forma independiente en múltiples animales paralelamente. Teniendo
eso en cuenta, es necesario considerar que dado que también se han encontrado
dinosaurios ornitísquios con cierto tipo de plumaje, es probable que esta
característica estuviese presente en los dinosaurios más primitivos y quizá,
incluso en el ancestro común de todos los dinosaurios, lo que da lugar a la
posibilidad de que todos los dinosaurios (y como tal, todo tipo de terópodos y no
exclusivamente, los coelurosaurios) pudiesen haber presentado esta
característica. No obstante, algunas muestras de piel recuperadas sugieren que
este no necesariamente fue el caso, aunque tampoco lleva a desconsiderar ni a
descartar esta posibilidad.

Velociraptor como lo conocemos hoy.
Ilustración de Matthew Martyniuk
2) El intelecto de los terópodos extintos más listos NO era
superior al de un ave
En un principio, se creía que los dinosaurios eran animales
torpes y poco complejos con una capacidad muy limitada para interactuar entre
sí y desempeñarse en su cambiante entorno. Hoy, sin embargo, ocurre lo opuesto.
Existe la creencia popular de que ciertos dinosaurios y específicamente,
algunos tipos de terópodos, como los dromaeosaurios poseían un nivel de
inteligencia extremadamente desarrollado. Dicha creencia normalmente se asocia
con la errónea representación de los Velociraptores de la saga "Jurassic
Park", donde se muestran con un nivel intelectual exagerado y comparable
al de varios mamíferos. Si bien es cierto que se ha determinado que muchos
terópodos avanzados tenían un intelecto superior al de otros tipos de
dinosaurios y que probablemente eran capaces de interactuar coordinadamente
unos con otros e incluso, de elaborar estrategias de caza eficaces, es de tener
en cuenta que éstos probablemente no eran más listos que algunas aves actuales.
Comparando el tamaño del cerebro de los dinosaurios con relación al de su
cuerpo, los científicos estiman que la inteligencia de algunos terópodos era
similar a la de los cocodrilos (los cuales responden de forma impulsiva a su
entorno) y el algunos casos, a la de muchas aves actuales (que pueden responder
a su entorno de una forma más calculadora, aunque no de un modo tan complejo
como se ve en muchos mamíferos), no siendo tan listos como para desarrollar un
señalado sentido del honor, realizar hazañas determinadas para superar ciertos
tipos de obstáculos como abrir puertas o efectuar estrategias altamente
complejas como tender trampas con mediante la utilización de cebos, aunque
también es cierto que tampoco eran tan torpes como se creía con anterioridad. Y
es que los terópodos, como todo animal, sólo tenían que ser lo suficientemente
listos para desempeñarse eficazmente en su ecosistema (por ejemplo, en el caso
de los terópodos carnívoros, éstos sólo debían ser marginalmente más listos que
sus presas para poder atraparlas).

Jinfengopteryx utilizando una herramienta para atrapar
insectos, imagen con alto contenido especulativo, pero dentro de los límites de
lo plausible.
Ilustración de H. Esdaile
3) Los terópodos no podrían abrir puertas
Aún si los terópodos contaran con la inteligencia suficiente
para superar ciertos obstáculos como puertas cerradas, éstos probablemente no
serían capaces de abrir puertas como lo hacían los ya mencionados Velociraptores
de "Jurassic Park". Esto se debe a que el movimiento de sus manos era
relativamente limitado. Además, contrario a la forma en que se presentan en los
medios, los terópodos no podían colocar sus manos en pronación, sino qué éstas
estaban posicionadas en supinación. Es decir, sus manos no podían posicionarse
de modo que las palmas apuntaran hacia el suelo o hacia atrás, sino de tal
manera que una apuntaba en la dirección de la otra. En los primates, la
pronación es realizable por el movimiento del radio con respecto al cúbito (los
dos huesos del antebrazo). En los terópodos, sin embargo, el extremo del radio más
cercano al codo estaba sujeto al cúbito, limitando la movilidad. El movimiento
en la muñeca también fue limitado en muchas especies, obligando a todo el
antebrazo y a la mano a moverse como una sola cosa, dándole poca flexibilidad y
haciendo que el único modo para que las manos pudiesen ser posicionadas en
pronación fuese ensanchando lateralmente las extremidades, como hacen las aves al
elevar sus alas. No obstante, es sabido que los terópodos más primitivos
presentaban un mayor grado de movilidad en las patas delanteras y hay pruebas
de que algunas especies más avanzadas retuvieron una estructura más flexible.
Aún así, esto no los dotaba de capacidades que requerirían una rotación
prominente de las extremidades (como lo sería abrir puertas, por ejemplo), ya
que no necesitaban realizar ese tipo actividad y por consiguiente, no debían
contar con ese tipo de adaptación. Los brazos de los terópodos estaban
perfectamente diseñados para el agarre y la manipulación de alimento y en
algunos casos, para trepar árboles y quizá, para cavar, actividades necesarias
para su supervivencia.
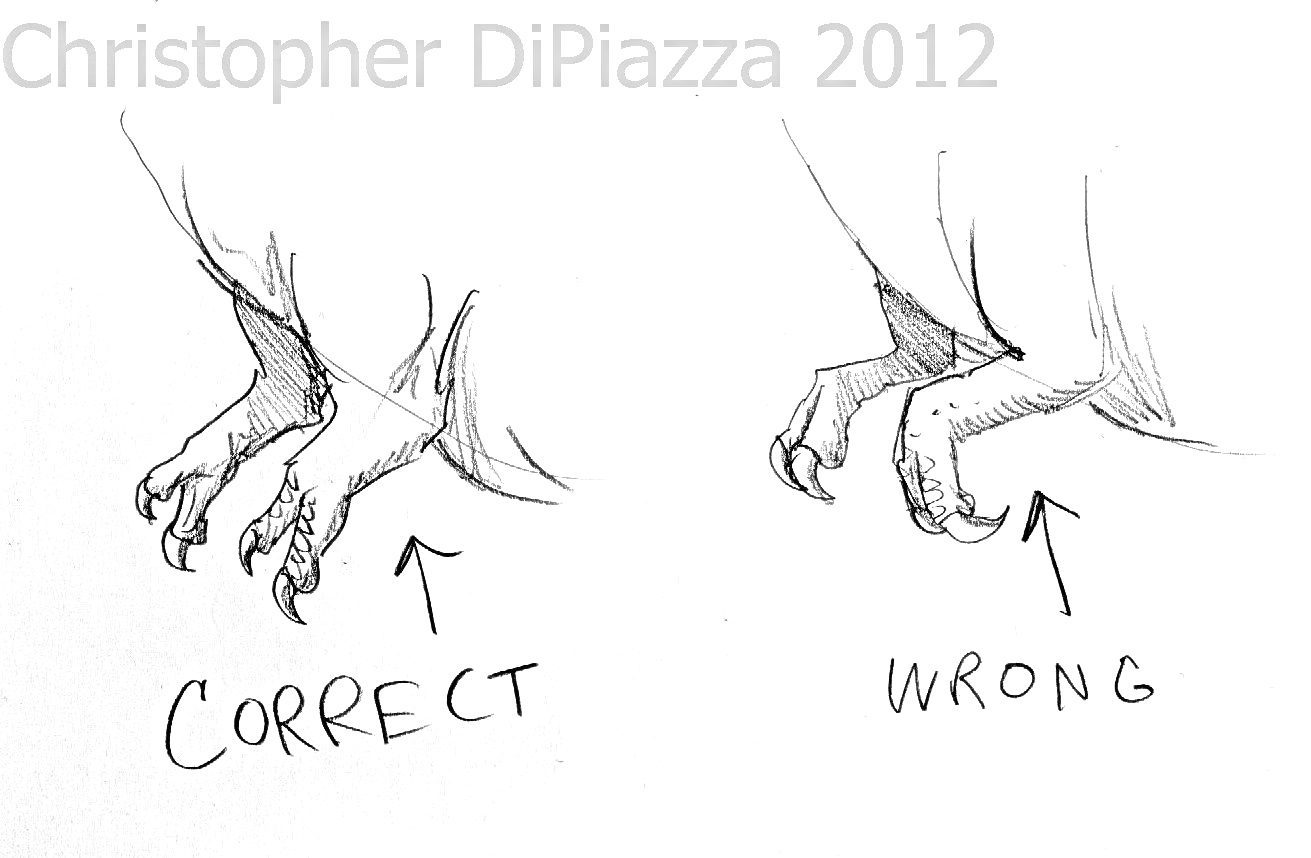
Comparación entre una versión errónea de un tiranosáurido
con sus manos en pronación y una versión correcta con manos en supinación
Ilustración de Christopher DiPiazza
4) Los terópodos quizá NO rugían como se suele pensar
Durante años, los medios han alimentado nuestra imaginación
y nuestros temores mostrándonos a los dinosaurios carnívoros emitiendo
aterradores rugidos con efectos de sonido muy bien trabajados para ese
propósito y hoy, estamos tan acostumbrados a eso que hasta se suele pensar que
es natural que estos animales hayan rugido de forma similar. La realidad, sin
embargo, pudo haber sido distinta. Hoy, los mamíferos emiten toda clase de
sonidos (incluyendo rugidos en algunos casos) gracias a un órgano conocido como
la laringe, el cual contiene las cuerdas vocales. No obstante, en los parientes
vivos de los dinosaurios, conocidos como los arcosaurios (grupo que contiene a los
pterosaurios, a los cocodrilianos y a los propios dinosaurios, incluyendo a las
aves), este órgano no contiene cuerdas vocales. En su lugar, las aves en este
caso, emiten sonidos gracias a otro tipo de órgano conocido como la siringe, el
cual depende de un sistema de sacos aéreos. Es sabido que los terópodos no
avianos también poseían un sistema de sacos aéreos, mas es de tener en cuenta
que éste, a diferencia del de las aves, no parece estar asociado a la
vocalización debido a que no hay registro de un saco clavicular, sin el cual,
la siringe (si es que la poseían) no funcionaría para emitir sonidos como en el
caso de las aves. Sin embargo, es de tener en cuenta que los dinosaurios han
sido uno de los grupos de animales más exitosos del planeta, lo cual está
interconectado con una capacidad de comunicación efectiva. En adición, se sabe
que los dinosaurios tienen un sistema auditivo desarrollado y que en algunos
terópodos, éste es bastante sofisticado. Además, es de considerar el hecho de
que algunas aves, como los buitres, no poseen siringe y de que en los
cocodrilianos, la laringe tampoco contiene cuerdas vocales y aún así, tanto los
buitres como los cocodrilianos pueden emitir sonidos controlando el flujo de
aire en sus sacos aéreos. Teniendo esto en cuenta, no es descabellado pensar
que al menos, algunos terópodos pudieran haber emitido alguna clase de sonido,
pero aún si este fuera el caso, el mismo no consistiría en rugidos, sino tal
vez en ronquidos o siseos de forma similar a los cocodrilianos, en graznidos
semejantes al de los buitres o en otros tipos de sonido que sólo requieran
flujo de aire en lugar de vocalización. No obstante, también está la
posibilidad de que los terópodos (y los dinosaurios no avianos en general)
contaran con un saco clavicular que simplemente, no se ha encontrado o que no
se preservó durante la fosilización o de que poseyeran algún otro tipo de órgano fónico o bien, de que su laringe fuese más
sofisticada y contara con la presencia de cuerdas vocales, pasando eventualmente a ser una característica vestigial, pero al no haber suficientes pruebas de que
alguno de éstos haya sido el caso, esta línea de pensamiento es difícil de confirmar
y enfrenta oposición. Si
bien es cierto que en películas como "Jurassic Park III" se alude a la comunicación de los dinosaurios e incluso, se menciona el
hallazgo de una cámara de resonancia de Velociraptor, el problema en realidad consiste
en que no hay pruebas directas que ofrezcan pistas sólidas sobre cómo se comunicaban los
dinosaurios.

Es posible que el icónico rugido del Tiranosaurio de
Jurassic Park no haya sido nada cercano al sonido que hacía el animal real.
Imagen de Universal Pictures
5) Es muy poco lo que sabemos a ciencia cierta sobre los
terópodos extintos
A cada rato, los terópodos nos sorprenden, mostrando ser un
grupo cada vez más complejo con cada descubrimiento. Y es que la paleontología,
como ciencia, con poca frecuencia ofrece respuestas certeras que puedan
considerarse como hechos irrefutables, abriendo así paso a múltiples
posibilidades y alternativas a considerar, sustituyendo conocimientos previos
al actualizarse con nuevos y corrigiendo percepciones que antes se consideraban
correctas, pero que hoy resultan no serlo. Por esta razón, los paleontólogos
constantemente dan a conocer información muy distinta a lo que creíamos saber. Ejemplo
de esto es el aspecto de dinosaurios de los que no poseíamos demasiado material
para estudiar (como ocurrió recientemente con el Espinosaurio y el Deinocheirus)
y como éstos, puede haber muchos casos, pues la realidad es que el estudio de
fragmentos de huesos, huellas y otros tipos de fósiles no es muy revelador
sobre cómo fue, por ejemplo, el comportamiento, las estrategias de caza, los
patrones de reproducción y en una gran mayoría de los casos, ni siquiera sobre
aspectos tan básicos como la apariencia física de criaturas que han estado
extintas durante millones de años. Gran parte de eso consiste mayormente en
deducciones y es producto de la especulación.

Ilustración de prehistoric-wildlife.com
Sin embargo, algo que sabemos con seguridad es
que los terópodos no son asesinos sádicos y siempre hambrientos, como se los
tiende a presentar en los medios. En la naturaleza, todo organismo hace lo que
es necesario para su supervivencia y sabemos que los terópodos no son la
excepción. Éstos han habitado el planeta durante más de 160 millones de años,
lo que significa que son un grupo sumamente exitoso, indicando a su vez que son
más complejos de lo que se suele pensar y que en lugar de ser criaturas irracionales
interesadas sólo en cazar y pelear con todo lo que se cruzara en su camino, la
ciencia indica que, como todo animal, han sido capaces de desempeñarse
eficazmente en su entorno, adaptándose para cumplir roles determinados en su
ecosistema y siendo cada especie eficiente y exitosa a su manera y capaz de
actuar en pro de su supervivencia, cosa que han hecho durante más de 225
millones de años y así lo sigue demostrando el único linaje de terópodos que ha
prevalecido hasta hoy, las aves.




























.jpg)





.jpg)







